
| 时间:2024-07-26 |
2023年发表于《PLoS Biology》(IF=7.8)
一、研究背景
先天性脑积水(CH)是一种常见的出生缺陷,其手术治疗有许多并发症,使CH成为儿童发病的重要原因。在Trim71基因中发现了许多CH相关的错义突变。Trim71是一种进化保守的RNA结合蛋白(RBP),在胚胎和神经干细胞中大量表达,Trim71对早期胚胎发生和神经正常分化至关重要。在机制上,Trim71结合靶mRNA并触发翻译抑制和/或mRNA降解。Trim71中CH相关突变导致干细胞分化和神经发生缺陷的分子机制仍不完全清楚。
本研究发现R783H-Trim71和R595H-Trim71导致小鼠胚胎干细胞(mESCs)的过早分化和加速神经谱系承诺,但每个Trim71突变体结合一个不同的异位靶mRNA亚群。发现R595H-Trim71特异性结合并抑制编码β-catenin的Ctnnb1 mRNA的翻译,导致Wnt/β-catenin信号活性降低,特别是在R595H-Trim71突变mESCs中。升高β-catenin水平可以缓解R595H-Trim71 mESCs的分化缺陷,而对R783H mESCs则没有作用。这些数据表明,Trim71 RNA结合域的功能获得性突变通过不同的病理机制导致CH,CH患者可以根据这些不同的机制进行分层,并且可以针对CH的遗传多样性病因设计精确的治疗策略。
二、研究结果
1、Trim71中两个与CH相关的突变导致了mESCs中类似的干细胞分化缺陷
使用mESCs作为Trim71 CH相关突变的机制研究模型,因为Trim71突变引起的发病机制在小鼠和人之间是保守的。使用CRISPR/cas9介导的基因组编辑将R595H或R783H引入Trim71中,获得了4个突变的mESC系,分别代表单等位基因(R595H/+,R783H/+)和双等位基因(R595H,R783H)突变。在Trim71突变体和WT mESCs中,多能因子的蛋白水平(图1A)和集落形成相似,表明自我更新不受突变的影响。然而,所有4个突变系都表现出分化缺陷,所有Trim71突变的mESCs都比WT型mESCs更快地失去了多能性(图1B);所有Trim71突变的mESCs都显示出较低水平的多能性因子,如Nanog(图1C-1E)。然而,与Trim71敲除(KO)mESCs相比,突变mESCs与WT mESCs具有相似的增殖和凋亡率。因此,Trim71突变mESCs的自发分化缺陷不是由增殖或凋亡的改变引起的。鉴于杂合子和纯合子突变体表现出相似的细胞和分子表型(图1A-1E),使用纯合子突变体mESCs进行机制研究。
研究发现,与WTmESCs相比,R595H-Trim71和R783H-Trim71mESCs的神经谱系承诺均增强,Sox1和Pax6的表达更高(图1F和1G),Sox1和Pax6是神经外胚层分化所必需的两种谱系特异性转录因子,这与最近的研究一致。两种Trim71突变mESCs的神经分化过程中,神经干/祖细胞标记物Nestin的表达也更高(图1H)。这些结果表明,Trim71中CH相关突变的mESCs更容易发生神经分化。
总之,Trim71中两种不同的CH相关突变导致胚胎干细胞分化和神经谱系承诺的相似缺陷。这些CH相关突变是功能获得突变,导致分化表型。

图1 Trim71中的两个CH相关突变在mESCs中引起类似的分化缺陷
2、这两个CH相关的Trim71突变体结合不同的底物mRNA
这两个CH相关突变都位于Trim71的RNA结合区域,为了确定R595H-Trim71是否结合了不同的mRNA底物库,通过交联免疫沉淀和测序(CLIP-seq)鉴定了转录组范围的靶标,并与R783H-Trim71和WT-Trim71的CLIP-seq数据进行了比较分析(图2A)。与随机序列相比,WT-Trim71和两个Trim71突变体的结合位点具有类似的预测茎环结构的过度代表性,但没有丰富的初级序列基序(图2C)。这些数据与Trim71识别结构基序的体外研究结果一致。与R783H突变类似,R595H突变也改变了Trim71的mRNA底物(图2B)。
两个Trim71突变体结合不同的mRNA组,重叠非常小(图2B)。我们通过CLIP-qRT-PCR验证了这一点(图2D和2E)。例如,R783H Trim71的靶标Lsd1 mRNA与R783H-Trim71相关,但与WT-或R595H-Trim71无关(图2E)。这些结果表明,不同的Trim71突变体结合不同的靶mRNA。
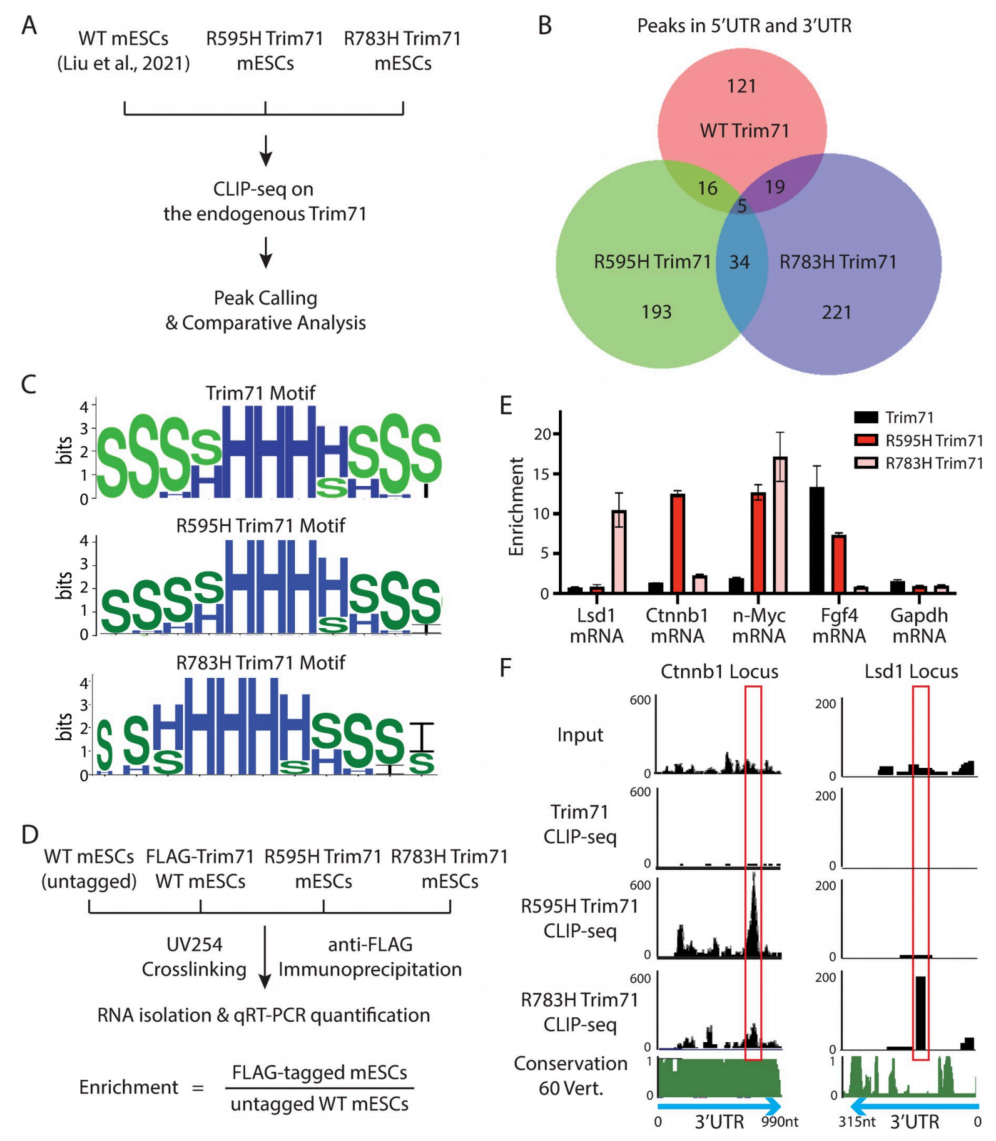
图2 WT Trim71和两个Trim71突变体在mESCs中转录组范围内的靶mRNA的比较分析
3、R595H-Trim71潜在功能靶标mRNA的鉴定
为了识别Trim71突变体的功能mRNA靶点,我们最初关注Trim71突变体结合的mRNA,因为两个突变体mESCs表现出相似的分化缺陷,而Trim71-KOmESCs不明显。在这34个共同的靶点中,重点关注了在mESCs中表达水平合理的靶点,以及在干细胞生物学和神经分化中具有注释功能的编码基因(图3A和3B)。在WT和两个Trim71突变体mESCs中,没有一个基因的蛋白水平显示出一致的变化(图3C)。这些结果表明,两个Trim71突变体在这些mRNA上的异位结合并不影响转录本的稳定性或翻译,并表明这些共同的靶点mRNA都不是Trim71突变体的功能靶点。
鉴于CH相关突变似乎具有不同的功能获得(图2B),因此考虑两种突变mESCs中类似缺陷可能是由结合和失调不同的靶mRNA引起的。最近发现Lsd1 mRNA是R783H-Trim71的功能靶标。本研究观察到LSD1蛋白在R783H mESCs异性降低,而在R595H mESCs中没有改变(图3C)。这一结果进一步支持了两个Trim71突变体的分子机制可能不同的说法。因此,为了鉴定R595H-Trim71的功能靶mRNA,对R595H-Trim71特异性结合的靶mRNA应用了类似的原理(图3A)。在我们分析的五个候选基因中,只有CTNNB1在R595H mESCs的蛋白水平上表现出一致和特异性的下降(图3B和3C),这表明Ctnnb1 mRNA是R595H-trim71的功能靶点。Ctnnb1 mRNA编码的β-catenin是经典Wnt/β-catenin信号通路的关键组成部分,对干细胞的正常分化至关重要。有趣的是,在Ctnnb1 mRNA的3’UTR中有一个主要的R595H-Trim71特异性结合位点(图2F)。此外,与许多mRNA不同的是,Ctnnb1 mRNA 30 UTR在脊椎动物中高度保守(图2F)。

图3 候选基因的功能鉴定
4、R595H-Trim71结合Ctnnb1 mRNA并抑制其翻译
多项证据表明,R595H-Trim71特异性抑制Ctnnb1 mRNA的翻译。首先,R595H-Trim71 mESCs特异性显示β-catenin水平降低,而Ctnnb1 mRNA水平没有变化(图4A-4C)。其次,Ctnnb1 mRNA与多体的关联在R595H-Trim71 mESCs异性降低(图4D和4E)。第三,R595H-Trim71在WT mESCs中的异位表达降低了β-catenin水平,但不降低Ctnnb1 mRNA水平(图4H-4J)。相比之下,R783H-Trim71特异性影响Lsd1 mRNA的蛋白水平和多体关联,而R595H-Trim71不受影响(图4A-4J)。总的来说,这些数据表明两个CH相关的Trim71突变体抑制不同的异位mRNA的翻译,R595H-Trim71特异性地抑制Ctnnb1 mRNA的翻译。因此,与R783H-Trim71对Lsd1 mRNA的调控类似,R595H-Trim71介导的Ctnnb1 mRNA的调控在小鼠和人之间是保守的。
总的来说,这些结果表明不同的Trim71突变体调节不同的异位靶mRNA翻译。
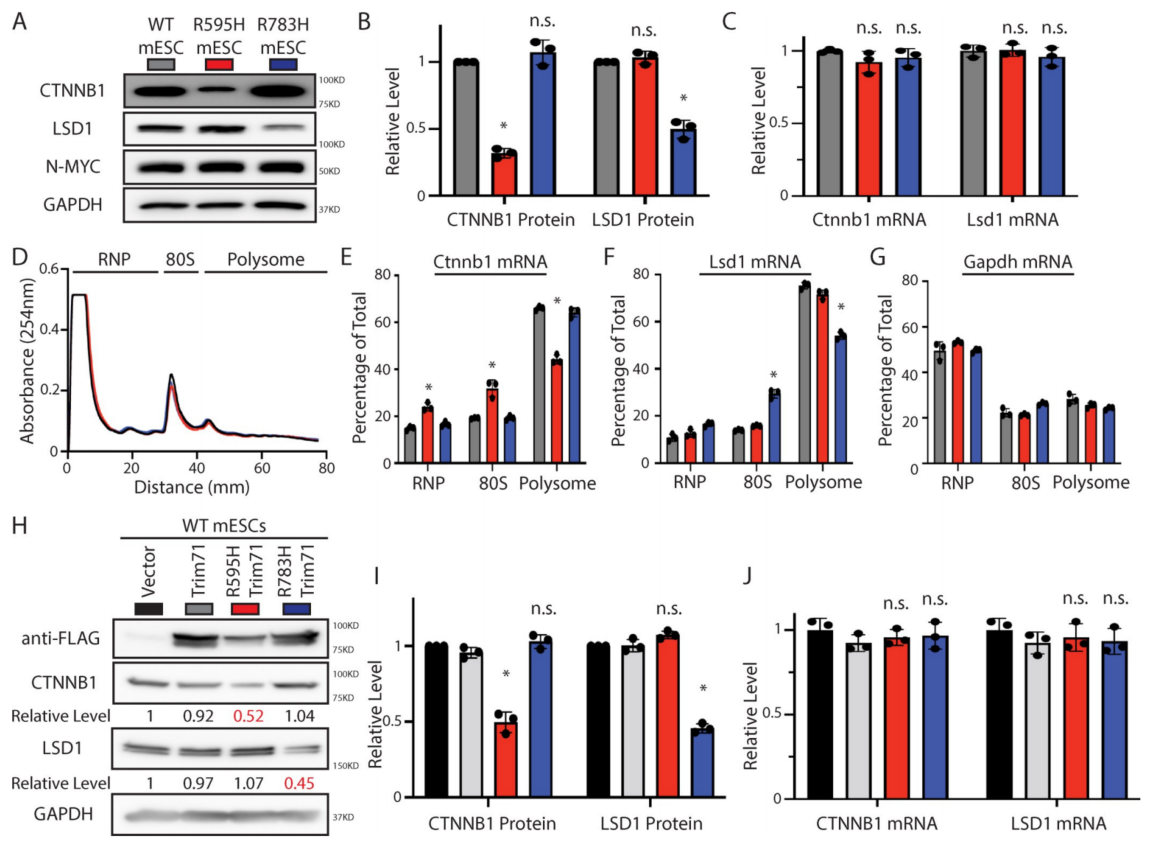
图4 这两个Trim71突变体抑制了mESCs中不同mRNA的翻译
5、消除异常R595H-Trim71:Ctnnb1 mRNA的相互作用可减轻mESC的分化缺陷
由于β-catenin是Wnt信号通路的关键成分,确定了β-catenin水平降低是否会影响R595H-Trim71 mESCs中的Wnt信号通路。使用荧光素酶报告基因来测量Wnt信号活性,其表达受Wnt信号响应元件(TCF/LEF)控制(图5A)。与WT和R783H-Trim71 mESCs相比,R595H-Trim71 mESCs中的Wnt信号活性降低(图5A),这表明Ctnnb1 mRNA是R595H-Trim71的功能相关靶标。为了确定Ctnnb1 mRNA的失调是否导致了R595H-Trim71 mESCs的分化缺陷,敲除了Ctnnb1 mRNA 3’UTR中的R595H-Trim71结合位点(称为“Ctnnb1 CLIPΔ”;图5B)。通过CLIP-qRT-PCR证实Ctnnb1-CLIPΔ特异性地破坏了Ctnnb1 mRNA与R595H-Trim71之间的相互作用(图5C)。重要的是,在表达Ctnnb1 CLIPΔ的R595H-Trim71 mESC中,β-catenin水平和Wnt信号活性得到恢复,而Ctnnb1 mRNA水平没有明显改变(图5D-5F)。这种β-catenin水平和Wnt信号的增加是R595H-Trim71 mESCs特有的,而在WT mESCs中表达Ctnnb1 CLIPΔ时没有观察到(图5D-5F)。
接下来,为了确定Ctnnb1 CLIPΔ是否也能缓解R595H-Trim71 mESCs的干细胞和神经分化缺陷,进行了出口多能性实验,并检测了多能性因子在分化过程中的表达。事实上,在表达Ctnnb1 CLIPΔ的R595H-Trim71 mESCs中,分化过程中的多能性和Nanog水平恢复了(图5G和5H);神经谱系规范因子Pax6和Sox1的水平在定向神经分化过程中得到恢复(图5I)。相比之下,Ctnnb1 CLIPΔ并没有减轻R783H-Trim71 mESCs的分化缺陷。此外,Lsd1 CLIPΔ虽然减轻了R783H-Trim71 mESCs的分化缺陷,但未能挽救R595H-Trim71 mESCs的缺陷。因此,每个CH相关的Trim71突变对分化的影响是靶mRNA特异性的。
综上结果表明,Ctnnb1 mRNA是R595H-Trim71的重要异位和功能靶点,Wnt/β-catenin信号通路的减少是R595H-Trim71 mESCs分化缺陷的主要和特异性原因。
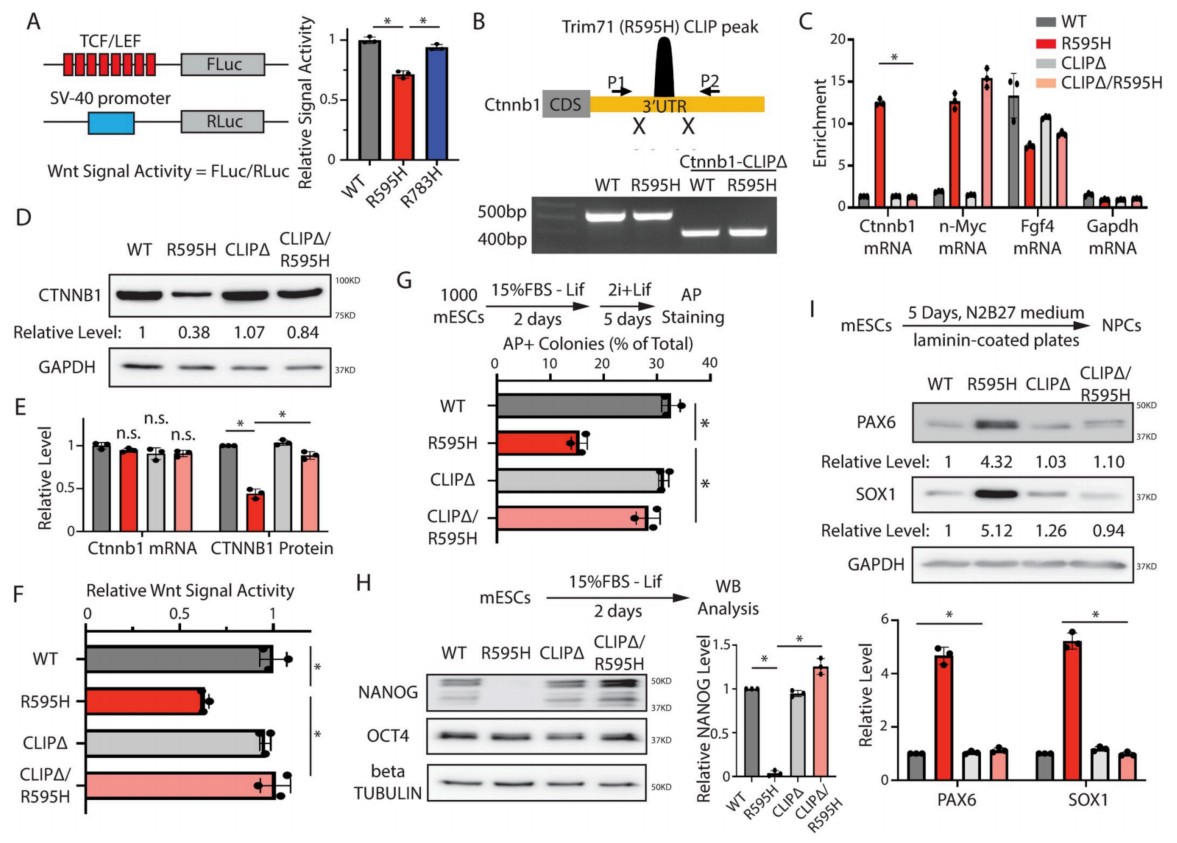
图5 破坏Ctnnb1 mRNA与R595H Trim71之间的相互作用,可特异性地缓解R595H mESCs的分化缺陷
6、增加β-catenin蛋白水平可特异性缓解R595H-Trim71mESCs的分化缺陷
基于目前的研究结果,推测增加β-catenin蛋白水平可以缓解R595H-Trim71 mESCs的分化缺陷。为了验证这一假设,用表征良好的Wnt信号激活剂BML-284处理R595H-Trim71 mESCs,以稳定β-catenin。发现BML-284处理的R595H-Trim71 mESCs,显示出分化缺陷的剂量依赖性缓解(图6A)。在R595H-Trim71自发性分化过程中,使用这种激动剂也能恢复Nanog水平,但在R783H-Trim71 mESCs中则没有恢复(图6B)。但在BML-284存在的情况下,观察到WT和突变mESCs在定向神经分化过程中显著的细胞死亡,这表明Wnt信号的组成性激活是有害的。作为一种正交的方法,通过异位表达Ctnn1b-GFP来增加β-catenin蛋白的水平(图6C)。β-catenin蛋白水平的升高减轻了R595H-Trim71mESCs细胞的分化缺陷(图6D、6E)。此外,在定向神经分化过程中,β-catenin蛋白水平的增加对WT mESCs没有影响,但减轻了R595H-Trim71 mESCs的过早神经谱系承诺(图6F)。
总之,Wnt/β-连环蛋白信号通路的降低特异性地导致了R595H-Trim71mESCs的干细胞分化缺陷。
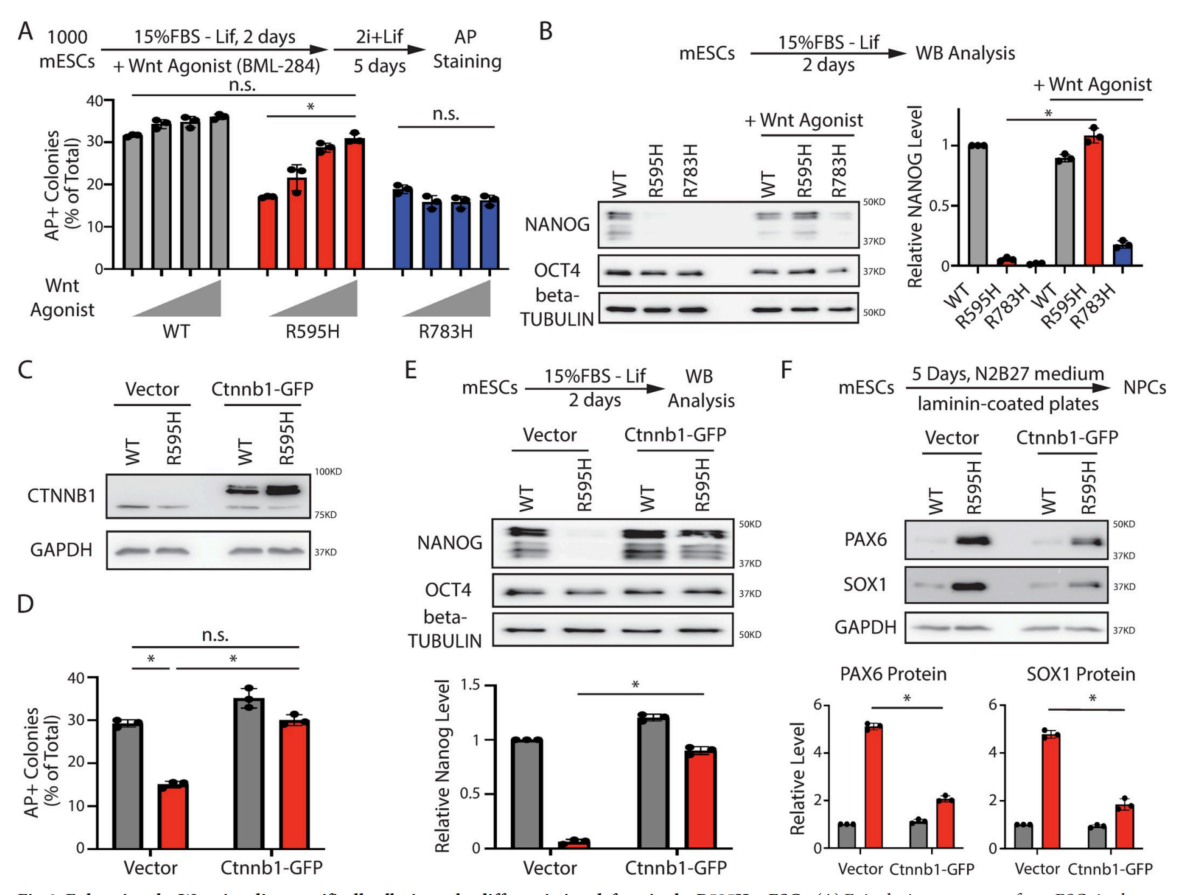
图6 增强Wnt信号可特异性地缓解R595HmESCs的分化缺陷
7、R595H-Trim71 mESCs中表达降低的β-catenin蛋白靶基因的鉴定
为了鉴定R595H-Trim71 mESCs中受Wnt/β-catenin蛋白信号通路减少影响的靶基因,测定了WT和R595H-Trim71 mESCs中与内源性β-catenin蛋白相关的染色质区域。β-catenin蛋白并不直接结合DNA/染色质,而是通过作为募集其他转录因子(如TCF和LEF)的支架来调节靶基因的表达。使用Cut&Run试验来确定目标位点(图7A),结果显示β-catenin相关染色质区域(峰)在WT mESCs的启动子区域周围富集(图7B)。此外,在Wnt信号通路的多个特征明确的靶基因中观察到β-catenin峰,如Sp6。
将来自WT的β-catenin峰与来自R595H-Trim71 mESCs的峰进行比较,发现它们在R595H-Trim71 mESCs中的强度整体下降(图7B和7C),与Wnt信号活性减弱一致。差异分析显示,R595H-Trim71 mESCs中有68个染色质区域,包括9个启动子,β-catenin关联显著减少(图7D),表明这些基因可能失调。其中Ralb在R595H-Trim71细胞中,其启动子区域的β-catenin峰显著降低(图7E),并且在R595H-Trim71 mESCs分化过程中,与WT mESCs相比,Ralb mRNA显著降低(图7F)。为了确定Ralb的减少是否会导致干细胞分化缺陷,检测了Ralb杂合mESCs的多能性退出。与R595H-Trim71 mESCs类似,Ralb杂合mESCs也表现出加速退出多能性(图7G)。这些结果表明,由于Wnt/β-catenin信号传导减少而导致的Ralb表达减少,可能导致R595H-Trim71 mESCs的分化缺陷。
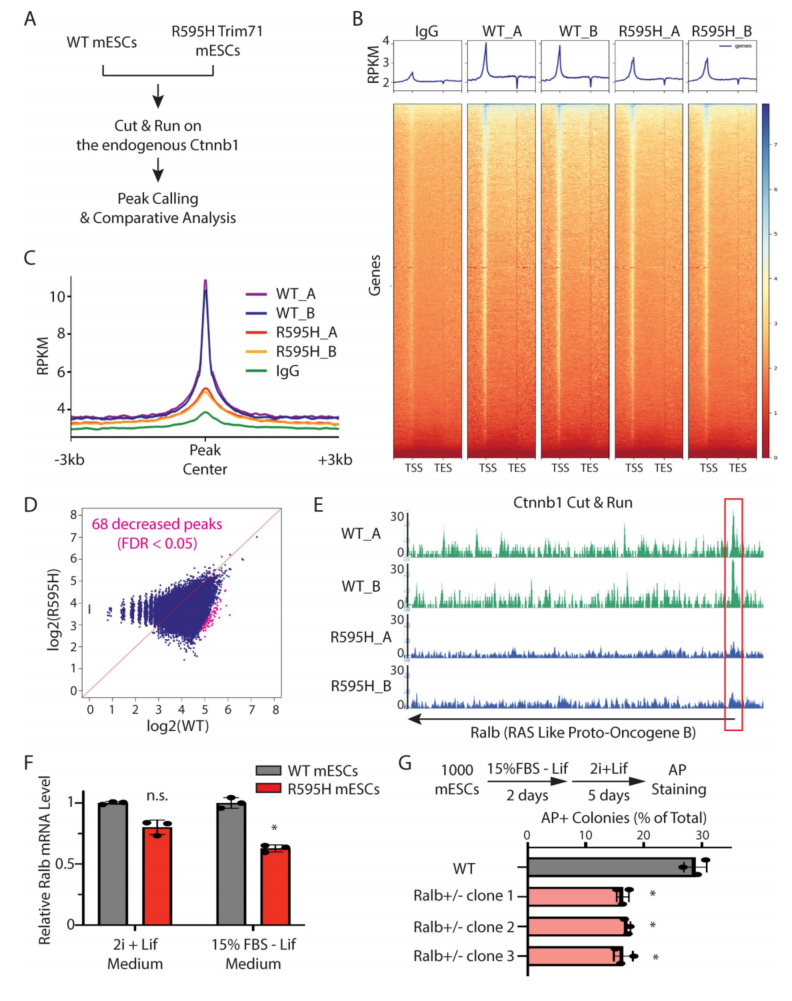
图7 鉴定R595H mESCs中受Wnt信号通路减少影响的靶基因
8、β-catenin蛋白的失调是具有单等位基因R595H-Trim71的mESCs分化缺陷的基础
为了评估双等位基因R595H mESCs机制是否具有病理学相关性,研究了Trim71中具有单等位基因R595H突变的mESCs(R595H/+),该突变模拟了人类CH的遗传突变模式。与纯合子R595H-Trim71 mESCs类似,与WT mESCs相比,杂合子R595H-Trim71 mESCs也具有较低的β-catenin水平(图8A),降低的Wnt信号活性(图8B)和增强的神经谱系承诺(图8C)。此外,β-catenin的异位表达(图8D)减轻了R595H-Trim71 mESCs的所有分化缺陷,如多能性退出率的恢复(图8E),自发分化过程中Nanog的表达的恢复(图8F),以及神经定向分化过程中神经谱系标记表达的恢复(图8G)。因此,在模拟CH的杂合环境中,升高β-catenin水平可以减轻Trim71中R595H突变引起的干细胞分化缺陷。
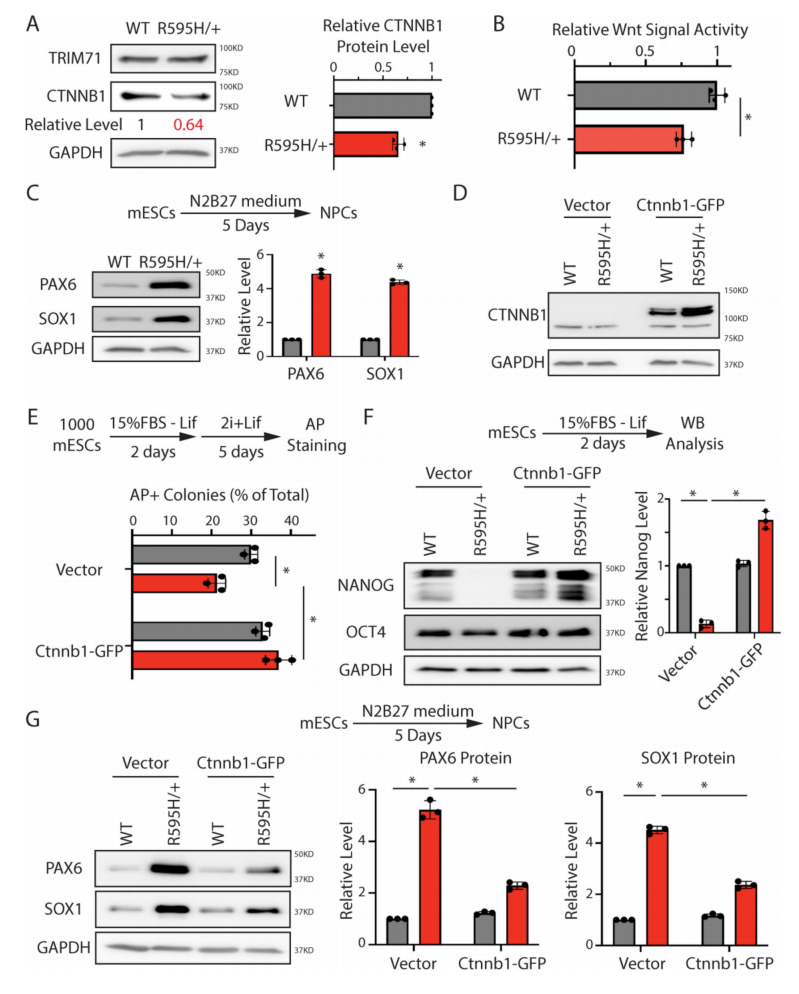
图8 下调的Ctnnb1对于Trim71中单等位基因R595H突变的mESCs的分化缺陷至关重要
三、研究结论
Trim71中两个与CH相关的突变体会结合不同的底物mRNA,R595H-Trim71结合Ctnnb1 mRNA并抑制其翻译,会引起β-catenin蛋白水平降低,从而导致Wnt/β-catenin信号传导减少,Ralb表达减少,引起R595H-Trim71 mESCs的分化缺陷。
参考文献:
Different congenital hydrocephalus–associated mutations in Trim71 impair stem cell differentiation via distinct gain-of-function meCHanisms.[J]PLoS Biology, 2023.